Bone Book Review Bone Book Review Robert Lubbers
Introduction
Crumbling may be a unique biological process, since evolutionarily at that place appears to be an absence of genes specifically selected to cause information technology (1, two). Rather, age-related changes may be the unprogrammed results of optimization for early on reproductive success. Thus, senescence at the organismal level represents a phenomenon with low mechanistic conservation among disparate metazoans and so mechanisms of human aging do not necessarily take metazoan counterparts in every situation. For example, replicative senescence or the loss of proliferative capacity in replication-competent somatic tissues is not a potential mechanism for aging in organisms where soma compartments are post-mitotic, such equally C. elegans (3).
A complementary arroyo to studying a model system for aging in a lower organism is to straight study human aging. Although this would closely capture aspects of aging that are relevant to humans, it does not obviate consideration for the highly polygenic nature of age-related pathologies (iv), the confounding effects of outbreeding, or environmental furnishings based on where and how individuals alive. An arroyo to providing a scientifically tractable system, at least with respect to the former, is to study genetic diseases whose phenotypes mimic at least some (i.eastward., "segmental") features of the usual human being aging procedure (4, 5). Such segmental progeroid (i.east., premature or accelerated aging-like) syndromes are unremarkably monogenic and may thus be simple enough to provide insights into the causes of their pathology. Studied within the context of theories for physiological aging, observations made in segmental progeroid syndromes may also explain sure aspects of normal crumbling. Despite being only fractional phenocopies of normal aging (i.due east., some tissues show aging features and other non), these segmental progeroid syndromes provide experimental tractability, with varying fidelity, that is the rationale for their use as epitome for natural deteriorative changes that occur over time. Single-gene mutations that touch on multiple aspects of the physiological crumbling phenotype may exert their activeness through developmental alterations that accept consequences for mail service-maturational crumbling, and importantly, for regulation of the rates of post-maturational aging later normal development.
Hither we propose that consideration of fibrodysplasia ossificans progressiva (FOP) as a segmental progeroid syndrome offers a unique perspective into potential mechanisms of normal aging and may also provide insight for identification of new targets for therapeutic interventions in FOP.
Segmental Progeroid Syndromes as a Model to Investigate Man Crumbling
Representative segmental progeroid syndromes are shown in Tabular array i. Several are monogenic or at least affect the aforementioned or like pathways when more than i factor causes the same phenotype within the same syndrome. The putative mechanism(s) by which aging phenotypes are manifested are like in several syndromes, including decreased genome maintenance and accelerated cellular senescence. All of the syndromes reduce mean lifespan or life expectancy.

Table 1. Representative segmental progeroid syndromes including FOP (4–7).
In the case of FOP, possible mechanisms for generation of an accelerated aging phenotype include injury-induced senescence and overactive activin A signaling. In comparison to other segmental progeroid syndromes, FOP represents an opportunity to study 2 different mechanisms by which crumbling phenotypes may be produced. Injury-induced senescence, particularly in soft tissue such as muscle, has recently been described (8, 9) and muscle injury is a known crusade of episodic inflammatory exacerbations or flare-ups in FOP (x, xi). Increased signaling through the bone morphogenetic protein (BMP) pathway, especially by activin A, has been implicated in osteoarthritis, sarcopenia, neurodegeneration, and other features associated with crumbling and FOP (discussed beneath under section Segmental Progeroid Features of FOP). Furthermore, activin A is a component of the senescence-associated secretory phenotype (SASP) (12, thirteen) and in FOP the mutated ACVR1/ALK2 encoding Activin A receptor type I/Activin-like kinase 2 (ACVR1/ALK2), a BMP blazon I receptor, is exquisitely sensitive to increased levels (14). Thus, injury-induced senescence leading to increased production of activin A may precipitate flare-ups in FOP and increased BMP signaling through activating mutations in ACVR1 may contribute to accelerated historic period-related changes in certain tissues. To exam this hypothesis it volition be necessary to examine the senescent cell burden in FOP lesion formation using markers of senescence in both patient samples and mouse models of FOP as well equally analysis of the SASP in mouse models of FOP.
Fibrodysplasia Ossificans Progressiva (FOP)
FOP is a strongly debilitating genetic disorder with hallmark features of congenital first toe malformations, progressive heterotopic ossification (HO) that produces normal bone at actress-skeletal locations, and accelerated features of crumbling (x, 11). The worldwide prevalence is 1/1,300,000–1/2,000,000 (15, 16). In that location is no ethnic, racial, gender, or geographic predilection to FOP. Early on in life, episodic bouts of inflammatory soft tissue protuberances (i.e., exacerbations or flare-ups) develop which are oft caused by injury, intramuscular injections, viral infections, muscular overuse, or fatigue (17, 18). These exacerbations convert connective tissues, including skeletal muscle, into HO. Tendons, ligaments, fascia, and aponeuroses are also afflicted, and together with transformed muscle, result in joint ankyloses and immobility. Singular forms of FOP have been reported (19). Approximately 97% of patients with FOP harbor an activating mutation (617G > A; R206H) in ACVR1/ALK2 (6). Individuals with FOP variants also take heterozygous ACVR1 missense mutations in conserved amino acids. FOP is diagnosed clinically, with confirmation by genetic testing if available. The majority of FOP cases are sporadic (i.due east., non-inherited mutations), but a small number of cases demonstrate germline transmission with inheritance in an autosomal dominant fashion (6). Although progressive HO is a hallmark characteristic, changes in early adulthood reminiscent of premature crumbling are evident.
Currently, there are no curative interventions, and the mainstay of treatment is focused on symptomatic relief using brief courses of loftier-dose corticosteroids for flare-ups, which may assistance to reduce the intense pain and edema associated with the early stages of ectopic bony lesion formation (10, 20). Steps to mitigate the likelihood of falls, pass up in pulmonary office, and acquisition of viral infections are important prophylactic measures. The median life expectancy is about 56 years of historic period (7). Most patients crave partial or complete help for ambulation by age 30, and common proximal causes of death include thoracic insufficiency syndrome and pneumonia (7). Factors contributing to the accelerated crumbling phenotype of FOP may be primarily related to ACVR1/ALK2 mutation, secondarily related to immobilization and disuse due to HO-associated joint ankyloses, or a combination of the two. Endpoints of current clinical trials focus on reducing heterotopic os formation (twenty), but it is unclear if those therapies targeting mutant ACVR1/ALK2 signaling will as well delay, prevent, or improve the progeroid features of FOP. Furthermore, it is unclear if targeting the activin-A ligand, vs. the receptor or post-receptor pathways, will be sufficient to mitigate all aspects of the condition. Also, information technology is unknown if or how targeting activin-A and its signaling networks will impact its role in hypothalamic-pituitary-gonadal feedback.
Segmental Progeroid Features of FOP
Progeroid features in FOP that may primarily exist associated with mutations in ACVR1 include alopecia, subcutaneous lipodystrophy, hearing loss, myelination defects, osteoarthritis, heightened inflammation, menstrual abnormalities, and maybe nephrolithiasis (Table two).

Tabular array 2. Progeroid features in FOP.
Alopecia is oftentimes observed in individuals with FOP and clinically is seen in both males and females. Evidence suggests that BMP signaling is involved in the control of the hair cycle (25). Increased BMP signaling through expression of BMP4, or its inhibition by the adversary Noggin, causes progressive baldness (26). In androgen-dependent alopecia, elevated BMP signaling in early (refractory) telogen likely mediates the retentivity of quiescent bulge stem cells (27). The case for elevated BMP signaling in lipodystrophy is less directly. Increased Fra-one causes severe lipodystrophy (21) and both BMP-2 and TGF-β stimulate AP-1 activities, including the DNA binding activity of Fra-1 (22). Alternative explanations for subcutaneous lipodystrophy include decreased caloric intake after jaw ankylosis and the effects of recurrent inflammatory flare-ups. With respect to the latter, activation of the NF-κβ pathway during periods of acute or chronic inflammation may contribute to loss of subcutaneous fat. For example, activation of the NF-κβ pathway due to ubiquitination defects has been associated with lipodystrophy (23, 24).
Conductive and sensorineural hearing loss are common in FOP (28) and with prebycusis. Conductive hearing loss occurs when sound waves are non relayed efficiently to the inner ear, while sensorineural hearing loss is related to sensory organ (cochlea and associated structures) dysfunction or damage to the vestibulocochlear nerve (cranial nerve Viii). In humans, NOGGIN (NOG) gene mutations are associated with a few autosomal dominant conditions similar proximal symphalangism and multiple synostoses which are characterized past skeletal defects and fusion of adjacent bones. Synostosis of one or more ossicles in the ear promotes conductive hearing loss. Proper formation of the skeleton requires balanced levels of BMPs and Noggin and the conductive hearing loss in Nog +/−mice results from an ectopic bridge of os between the stapes and the tympanum, interfering with the normal mobility of the ossicle (29). BMP signaling is likewise required for inner ear development, including patterning of sensory regions in the cochlea that process audio (30). Information technology is probable that hearing loss in FOP is due to increased BMP signaling very early in life affecting both/either the cochlear sensory regions and/or motion of ossicles. Later in life, synostosis of the ossicles due to HO may be the predominant cause of progressive hearing loss.
Demyelinated lesions and focal inflammatory changes of the CNS are seen in both mouse models of FOP and in CNS white matter lesions in FOP patients (31). BMP signaling is a stiff inhibitor of oligodendroglial differentiation and remyelination (32), and gain-of-function mutations in ACVR1/ALK2 predictably enhance this potent inhibition. Dysregulated BMP signaling causes CNS demyelination, and CNS demyelination is one of the underlying mechanisms for the observed singular neurologic phenotypes in FOP patients. With normal aging, decreased CNS remyelination becomes more prominent over fourth dimension (33, fifty). In addition, aging is associated with decreased hippocampal neurogenesis and concomitant hippocampus-dependent cerebral functions (51). There is an inverse human relationship between CNS levels of BMP4 expression and noggin with historic period, with the former increasing essentially in the mouse dentate gyrus and the latter decreasing. This results in a profound elevation of phosphorylated-SMAD1/5/8, a key effector of BMP signaling. Every bit with crumbling in mice, a large increase in BMP4 expression is seen in the dentate gyrus of older humans without known cognitive dysfunction (51). Increased BMP signaling is related to impairments in neurogenesis and to historic period-related cognitive changes (51) and aspects of these processes may be phenocopied in FOP.
Accelerated osteoarthritis is commonly found in FOP. Terminal differentiation of chondrocytes may exist delayed or prevented past absconding of BMP signaling in articular cartilage, and mitigation of this blockage or increased BMP signaling may so contribute to endochondral ossification and breakdown of cartilage matrix (39). In cartilage, TGFß and BMP are necessary for normal articulation evolution and maintenance and their dysregulation has been associated with the pathogenesis of osteoarthritis. Interestingly, osteoarthritic patients take significantly higher serum levels of BMP-2 and BMP-4 compared to non-diseased humans and appear to narrate patients who accept degenerative joint disease astringent enough to require total joint replacement (40).
Heightened inflammation in FOP can be acute (as in episodic flare-ups) besides as chronic (as in an elevated pro-inflammatory state). The inflammatory nature of flare-ups in FOP is clinically obvious and well-described (41). In FOP patients without clinically evident HO, increased serum levels of cytokines, including IL3-, IL-vii, IL-eight, and IL-10, suggest a persistent pro-inflammatory state (42). So-called "inflammaging" refers to the chronic, sterile, low-grade inflammation which develops as part of normal aging, and is thought to contribute to the pathogenesis of multiple age-related diseases (43). In FOP, both astute and chronic inflammation may be related to the role of activin A in the initiation and persistence of the inflammatory response (44, 45).
Early menstrual abnormalities in FOP, including amenorrhea, are clinically recognized but accept not been objectively studied or described. Roles for activin A in the ovulation cycle likewise every bit in endometrial repair after catamenia have been reported and are perhaps causally related (46, 47).
Progeroid features in FOP that may secondarily be related to immobilization from progressive HO include decreased vital chapters, osteoporosis, fractures, sarcopenia, and predisposition to respiratory infections (Tabular array ii). These manifestations represent an opportunity to report the contribution of decay to the normal crumbling phenotype typified by the decreased physical activity, sedentary predilection, and increased likelihood of prolonged bed residual in older adults. If physiological aging is the issue of main aging processes interacting with or superimposed upon the pathophysiological consequences of inactivity (36), then specific characteristics of precocious aging in FOP due to disuse would be amenable to written report in isolation. Every bit an illustration, unloading of the normotopic skeleton due to bridging heterotopic bone results in osteoporosis. Some other case is the increase in chest wall rigidity and decreases in rubberband recoil and force-generating capacity of respiratory muscles that contribute to diminished vital capacity and predisposition to respiratory infection.
Some manifestations of precocious crumbling may be attributed to both primary and secondary furnishings of FOP. Sarcopenia in FOP probable represents the effects of both disuse cloudburst due to articulation ankyloses as well as increased activin A signaling causing both increased muscle catabolism and inhibition of myoblast differentiation (37, 38). Nephrolithiasis in FOP could be related to inadequate fluid intake due to functional difficulties in voiding, immobilization itself, and the effects of activin A on kidney function (48, 49).
Injury, Reprogramming in vivo, and Cellular Senescence
Injury, in general, is associated with accumulation of senescent cells [see (8) and Figure 1]. Growing evidence suggests that injury-induced reprogramming in skeletal muscle is facilitated by the accumulation of senescent cells at or almost the site of damaged tissue (eight). Bothe acute and chronic injury enables transcription-factor-mediated reprogramming in damaged muscle (8). The reprogramming issue of senescence appears to exist due to the release of interleukin half dozen (IL-6) and perhaps other components of the senescence-associated secretory phenotype (SASP) (8, 52). Senescence and the SASP facilitate the reprogramming of neighboring not-senescent cells merely also recruit macrophages for the removal of necrotic tissue (8).

Figure one. Musculus injury-induced senescence. Senescence-associated β-galactosidase (SAβ-gal) staining of the tibialis anterior musculus of a wild-blazon mouse is shown without injury (left) and 5 days after injury via cardiotoxin (CTX) injection. SAβ-gal-stained cells appear bluish. Images are courtesy of Haitao Wang, Ph.D., Mayo Clinic, Rochester, MN, Usa).
Paracrine release of IL-6 and other factors secreted past senescent cells promote reprogramming by Oct4, Sox2, Klf4, and c-Myc (OSKM) in not-senescent cells (53) into pluripotent cells (as well called induced pluripotent stem cells or iPSCs). A direct human relationship has been demonstrated between senescence and OSKM-driven reprogramming. In cells defective p16INK4a/ARF (i.due east., cells that do non undergo senescence), their ability to reprogram is severely compromised (9, 53). Furthermore, pharmacological inhibition of NFkB, a major driver of cytokine production and the SASP, reduces in vivo reprogramming (nine). Aging, which is associated with higher levels of cellular senescence, likewise favors OSKM-driven reprogramming. Similarly, in physiological atmospheric condition of wound healing, senescence triggered past injury could promote cell dedifferentiation to mediate repair of damaged tissue (nine, 53).
Roles of Cellular Senescence in FOP Lesion Formation
In FOP, injuries due to soft tissue trauma, viral infection, muscular stretching, and even fatigue due to overuse tin can precipitate a flare-up. Tissue damage causes pathogen-associated molecular patterns (PAMPs) and impairment-associated molecular patterns (DAMPs) in response to microbial and endogenous injury in the setting of a hypoxic microenvironment (54, 55). Every bit the consequence of tissue injury, senescent cells accrue and potentially contribute to early events that raise BMP signaling and facilitate the reprogramming of tissue-resident stem cells (Figure 2).
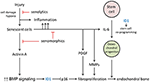
Figure two. Potential roles for cellular senescence in FOP lesion germination. The major hypothesized contributions of senescence are through the product of activin A, IL-6, and other components of the SASP. T, inhibitory pathways;, inhibitory activeness of senotherapeutic drugs.
Senescence is a cellular response to damage characterized past an irreversible cell cycle arrest and and then by the SASP (56, 57). The SASP produces at least two factors that can straight promote increased BMP signaling and stem cells reprogramming—activin A and IL-6, respectively (Figure ii). It is well-established that activin A stimulates BMP signaling in FOP cells, owing to the causative mutations in the ACVR1 gene. In addition to the permissive effects of IL-6 in reprogramming, FOP cells show an increased efficiency of iPSC generation (58). In normal cells, the generation of iPSCs is facilitated by transduction of mutant ACVR1 or SMAD1 or past the early on add-on of BMP4 during the reprogramming. ID genes, downstream targets of BMP-SMAD signaling, are important for iPSC generation and their signaling through this pathway can inhibit prison cell senescence due to p16/INK4A, which otherwise serves to prevent reprogramming (58). Thus, ID1 and other ID genes may serve to both enhance expansion of the FOP early lesion also as stimulate production of osteochondral progenitor cells. Enhanced BMP signaling promotes a tremendous fibroproliferative response, mayhap further accelerated past secretion of platelet-derived growth factor (PDGF) and matrix metalloproteinases (MMPs) by the SASP (Effigy 2). Osteochondral progenitor cells derived from reprogrammed stem cells ultimately contribute to the endochondral bone formation which is the hallmark of mature FOP lesions. Other events that may contribute to formation of heterotopic bone cannot be excluded (59). However, in mouse models it will exist possible to demonstrate if senescence-mediated tissue reprogramming in FOP lesions shifts lineage determination from a myogenic to a chondrogenic fate after injury.
Senescent cells may play multiple roles in the formation of HO in FOP and drugs which target senescent cells and/or the SASP may be candidates for therapeutic interventions. Compounds which selectively clear senescent cells (and then-called senolytics) were first described on the basis of targeting pro-survival networks in senescent cells (60). Compounds that reduce the SASP (i.eastward., senomodulators), including inhibitors of the JAK/STAT pathway that plays an important part in regulating cytokine production, reduce systemic and adipose tissue inflammation in old mice (61). Rapamycin, a senomodulator, may be of particular do good in FOP, since it likewise reduces activin-A mediated mTOR signaling (62). Many senotherapeutic agents take been reported, are effective in delaying or alleviating multiple age-related conditions in pre-clinical models, and are now being evaluated in clinical trials (63, 64). Their potential use in FOP offers a novel therapeutic arroyo to injury-induced flare-ups in FOP which should be farther explored. Nosotros propose that senescent prison cell clearance and/or reduction in the SASP will ameliorate HO formation in mouse models of FOP and can be translated for use in patients with FOP.
Conclusions
Monogenic segmental progeroid syndromes are important models for studying aspects of physiological aging. Features of precocious aging in FOP include both those that are related to dysregulated BMP signaling equally well as those secondary to early immobilization and decay. At the level of lesion formation in FOP, soft tissue injury resulting in hypoxia, cell harm, and inflammation may result in the accumulation of senescent cells as in aged tissue. Production of Activin A, interleukin vi, and other inflammatory cytokines as office of the SASP could mediate the initial signaling cascade that results in intense fibrosis too every bit tissue-resident stem cell reprogramming prior to ectopic endochondral bone formation. This proposal requires experimental validation, but is amendable to testing in animal models. Consideration of FOP as a segmental progeroid syndrome may offer a unique perspective into potential mechanisms of normal aging, may increment agreement of BMP signaling as related to os homeostasis and repair, and may too provide insight for identification of new targets for therapeutic interventions in FOP such every bit use of senotherapeutic drugs now in phase i and phase 2 clinical trials for aging-related conditions.
Author Contributions
RP conceived the work, with substantial contributions from HW and FK. RP drafted the initial manuscript with HW and FK revising it critically. RP, HW, and FK gave final approval to the work and agreed to exist answerable for all aspects.
Disharmonize of Interest
The authors declare that the research was conducted in the absence of whatsoever commercial or financial relationships that could be construed as a potential conflict of involvement.
Acknowledgments
This work was enabled by support by the Radiant Hope Foundation (to RP, HW), the Robert and Arlene Kogod Professorship in Geriatric Medicine (to RP), and the Isaac and Rose Nassau Professorship of Orthopedic Molecular Medicine (to FK).
References
iii. Herndon LA, Schmeissner PJ, Dudaronek JM, Brown PA, Listner KM, Sakano Y, et al. Stochastic and genetic factors influence tissue-specific decline in ageing C. elegans. Nature. (2002) 419:808–xiv. doi: x.1038/nature01135
PubMed Abstract | CrossRef Full Text | Google Scholar
5. Martin GM. Genetic syndromes in man with potential relevance to the pathobiology of aging. Birth Defects Orig Artic Ser. (1978) 14:v–39.
PubMed Abstruse | Google Scholar
half-dozen. Shore EM, Xu M, Feldman GJ, Fenstermacher DA, Cho TJ, Choi IH, et al. A recurrent mutation in the BMP blazon I receptor ACVR1 causes inherited and desultory fibrodysplasia ossificans progressiva. Nat Genet. (2006) 38:525–7. doi: 10.1038/ng1783
PubMed Abstruse | CrossRef Full Text | Google Scholar
7. Kaplan FS, Zasloff MA, Kitterman JA, Shore EM, Hong CC, Rocke DM. Early mortality and cardiorespiratory failure in patients with fibrodysplasia ossificans progressiva. J Bone Joint Surg Am. (2010) 92:686–91. doi: 10.2106/JBJS.I.00705
PubMed Abstract | CrossRef Full Text | Google Scholar
viii. Chiche A, Le Roux I, von Joest M, Sakai H, Aguin SB, Cazin C, et al. Injury-induced senescence enables in vivo reprogramming in skeletal muscle. Prison cell Stalk Prison cell. (2017) xx:407–14.e4. doi: ten.1016/j.stalk.2016.eleven.020
PubMed Abstruse | CrossRef Total Text | Google Scholar
9. Mosteiro 50, Pantoja C, Alcazar N, Marion RM, Chondronasiou D, Rovira Thou, et al. Tissue damage and senescence provide critical signals for cellular reprogramming in vivo. Science. (2016) 354:aaf4445. doi: x.1126/science.aaf4445
PubMed Abstruse | CrossRef Full Text | Google Scholar
eleven. Pignolo RJ, Shore EM, Kaplan FS. Fibrodysplasia ossificans progressiva: diagnosis, management, and therapeutic horizons. Pediatr Endocrinol Rev. (2013) 10(Suppl 2):437–48.
PubMed Abstract | Google Scholar
13. Xu M, Palmer AK, Ding H, Weivoda MM, Pirtskhalava T, White TA, et al. Targeting senescent cells enhances adipogenesis and metabolic part in old age. Elife. (2015) 4:e12997. doi: 10.7554/eLife.12997.028
PubMed Abstract | CrossRef Full Text | Google Scholar
xiv. Wang H, Shore EM, Pignolo RJ, Kaplan FS. Activin A amplifies dysregulated BMP signaling and induces chondro-osseous differentiation of chief connective tissue progenitor cells in patients with fibrodysplasia ossificans progressiva (FOP). Bone. (2018) 109:218–24. doi: 10.1016/j.bone.2017.eleven.014
PubMed Abstract | CrossRef Total Text | Google Scholar
15. Baujat Chiliad, Choquet R, Bouee Due south, Jeanbat Five, Courouve L, Ruel A, et al. Prevalence of fibrodysplasia ossificans progressiva (FOP) in French republic: an estimate based on a record linkage of ii national databases. Orphanet J Rare Dis. (2017) 12:123. doi: 10.1186/s13023-017-0674-5
PubMed Abstract | CrossRef Full Text | Google Scholar
17. Pignolo RJ, Bedford-Gay C, Liljesthrom Grand, Durbin-Johnson BP, Shore EM, Rocke DM, et al. The natural history of flare-ups in fibrodysplasia ossificans progressiva (FOP): a comprehensive global cess. J Bone Miner Res. (2016) 31:650–6. doi: 10.1002/jbmr.2728
PubMed Abstract | CrossRef Full Text | Google Scholar
19. Kaplan FS, Xu Thousand, Seemann P, Connor JM, Glaser DL, Carroll L, et al. Classic and atypical fibrodysplasia ossificans progressiva (FOP) phenotypes are caused by mutations in the bone morphogenetic protein (BMP) blazon I receptor ACVR1. Hum Mutat. (2009) xxx:379–90. doi: 10.1002/humu.20868
PubMed Abstract | CrossRef Full Text | Google Scholar
twenty. Kaplan FS, Pignolo RJ, Al Mukaddam MM, Shore EM. Hard targets for a second skeleton: therapeutic horizons for fibrodysplasia ossificans progressiva (FOP). Expert Opin Orphan Drugs. (2017) 5:291–4. doi: ten.1080/21678707.2017.1304211
PubMed Abstract | CrossRef Full Text | Google Scholar
21. Luther J, Driessler F, Megges One thousand, Hess A, Herbort B, Mandic V, et al. Elevated Fra-1 expression causes severe lipodystrophy. J Cell Sci. (2011) 124(Pt 9):1465–76. doi: 10.1242/jcs.079855
PubMed Abstruse | CrossRef Full Text | Google Scholar
22. Lai CF, Cheng SL. Signal transductions induced by bone morphogenetic protein-2 and transforming growth factor-beta in normal human osteoblastic cells. J Biol Chem. (2002) 277:15514–22. doi: ten.1074/jbc.M200794200
PubMed Abstract | CrossRef Full Text | Google Scholar
23. Aksentijevich I, Zhou Q. NF-κB pathway in autoinflammatory diseases: dysregulation of protein modifications by ubiquitin defines a new category of autoinflammatory diseases. Front Immunol. (2017) 8:399. doi: x.3389/fimmu.2017.00399
PubMed Abstract | CrossRef Total Text | Google Scholar
25. Rishikaysh P, Dev K, Diaz D, Qureshi WM, Filip Southward, Mokry J. Signaling involved in hair follicle morphogenesis and development. Int J Mol Sci. (2014) 15:1647–lxx. doi: ten.3390/ijms15011647
PubMed Abstract | CrossRef Full Text | Google Scholar
26. Botchkarev VA, Botchkareva NV, Roth W, Nakamura M, Chen LH, Herzog W, et al. Noggin is a mesenchymally derived stimulator of hair-follicle induction. Nat Cell Biol. (1999) ane:158–64. doi: 10.1038/11078
PubMed Abstract | CrossRef Full Text | Google Scholar
27. Nesterova A, Yuryev A. Androgenic alopecia: cross-talk betwixt cell signal transduction pathways. In: Pilus and Scalp Disorders. Intech (2017), p. 141–74. doi: x.5772/67845
CrossRef Full Text | Google Scholar
28. Levy CE, Lash AT, Janoff HB, Kaplan FS. Conductive hearing loss in individuals with fibrodysplasia ossificans progressiva. Am J Audiol. (1999) eight:29–33. doi: 10.1044/1059-0889(1999/011)
PubMed Abstract | CrossRef Total Text | Google Scholar
30. Ohyama T, Basch ML, Mishina Y, Lyons KM, Segil Northward, Groves AK. BMP signaling is necessary for patterning the sensory and nonsensory regions of the developing mammalian cochlea. J Neurosci. (2010) thirty:15044–51. doi: 10.1523/JNEUROSCI.3547-10.2010
PubMed Abstruse | CrossRef Full Text | Google Scholar
31. Kan L, Kitterman JA, Procissi D, Chakkalakal S, Peng CY, McGuire TL, et al. CNS demyelination in fibrodysplasia ossificans progressiva. J Neurol. (2012) 259:2644–55. doi: 10.1007/s00415-012-6563-x
PubMed Abstruse | CrossRef Total Text | Google Scholar
32. Sabo JK, Aumann TD, Merlo D, Kilpatrick TJ, Cate HS. Remyelination is altered by bone morphogenic protein signaling in demyelinated lesions. J Neurosci. (2011) 31:4504–10. doi: ten.1523/JNEUROSCI.5859-10.2011
PubMed Abstract | CrossRef Full Text | Google Scholar
33. Shields SA, Gilson JM, Blakemore WF, Franklin RJ. Remyelination occurs equally extensively merely more slowly in onetime rats compared to immature rats post-obit gliotoxin-induced CNS demyelination. Glia. (1999) 28:77–83. doi: 10.1002/(SICI)1098-1136(199910)28:1<77::AID-GLIA9>three.0.CO;2-F
PubMed Abstract | CrossRef Full Text | Google Scholar
34. Kussmaul WG, Esmail AN, Sagar Y, Ross J, Gregory S, Kaplan FS. Pulmonary and cardiac function in advanced fibrodysplasia ossificans progressiva. Clin Orthop Relat Res. (1998) 104–9. doi: 10.1097/00003086-199801000-00015
PubMed Abstruse | CrossRef Full Text | Google Scholar
35. Kaplan FS, Strear CM, Zasloff MA. Radiographic and scintigraphic features of modeling and remodeling in the heterotopic skeleton of patients who accept fibrodysplasia ossificans progressiva. Clin Orthop Relat Res. 1994:238–47. doi: ten.1097/00003086-199407000-00037
CrossRef Full Text | Google Scholar
37. Ding H, Zhang Grand, Sin KW, Liu Z, Lin RK, Li G, et al. Activin A induces skeletal muscle catabolism via p38beta mitogen-activated protein kinase. J Cachexia Sarcopenia Muscle. (2017) 8:202–12. doi: x.1002/jcsm.12145
PubMed Abstract | CrossRef Full Text | Google Scholar
38. Trendelenburg AU, Meyer A, Jacobi C, Feige JN, Glass DJ. TAK-1/p38/nNFκB signaling inhibits myoblast differentiation by increasing levels of Activin A. Skelet Muscle. (2012) 2:3. doi: 10.1186/2044-5040-2-3
PubMed Abstract | CrossRef Full Text | Google Scholar
39. Bauge C, Girard N, Lhuissier East, Bazille C, Boumediene G. Regulation and role of TGFβ signaling pathway in aging and osteoarthritis joints. Aging Dis. (2014) 5:394–405. doi: ten.14336/AD.2014.0500394
PubMed Abstract | CrossRef Full Text | Google Scholar
forty. Albilia JB, Tenenbaum HC, Clokie CM, Walt DR, Baker GI, Psutka DJ, et al. Serum levels of BMP-ii, 4, 7 and AHSG in patients with degenerative joint disease requiring total arthroplasty of the hip and temporomandibular joints. J Orthop Res. (2013) 31:44–52. doi: 10.1002/jor.22182
PubMed Abstract | CrossRef Full Text | Google Scholar
41. Convente MR, Wang H, Pignolo RJ, Kaplan FS, Shore EM. The immunological contribution to heterotopic ossification disorders. Curr Osteoporos Rep. (2015) xiii:116–24. doi: ten.1007/s11914-015-0258-z
PubMed Abstract | CrossRef Total Text | Google Scholar
42. Barruet E, Morales BM, Cain CJ, Ton AN, Wentworth KL, Chan TV, et al. NF-κB/MAPK activation underlies ACVR1-mediated inflammation in human heterotopic ossification. JCI Insight. (2018) three:122958. doi: ten.1172/jci.insight.122958
PubMed Abstract | CrossRef Full Text | Google Scholar
43. Franceschi C, Garagnani P, Parini P, Giuliani C, Santoro A. Inflammaging: a new immune-metabolic viewpoint for historic period-related diseases. Nat Rev Endocrinol. (2018) xiv:576–xc. doi: 10.1038/s41574-018-0059-4
PubMed Abstract | CrossRef Full Text | Google Scholar
44. Hedger MP, Winnall WR, Phillips DJ, de Kretser DM. The regulation and functions of activin and follistatin in inflammation and immunity. Vitam Horm. (2011) 85:255–97. doi: 10.1016/B978-0-12-385961-7.00013-5
PubMed Abstract | CrossRef Full Text | Google Scholar
45. Jones KL, Mansell A, Patella S, Scott BJ, Hedger MP, de Kretser DM, et al. Activin A is a disquisitional component of the inflammatory response, and its binding protein, follistatin, reduces mortality in endotoxemia. Proc Natl Acad Sci Usa. (2007) 104:16239–44. doi: 10.1073/pnas.0705971104
PubMed Abstract | CrossRef Full Text | Google Scholar
48. Maeshima A, Miya G, Mishima One thousand, Yamashita S, Kojima I, Nojima Y. Activin A: autocrine regulator of kidney development and repair. Endocr J. (2008) 55:i–9. doi: x.1507/endocrj.KR-113
PubMed Abstract | CrossRef Total Text | Google Scholar
49. Gupta RR, Delai PLR, Glaser DL, Rocke DM, Al Mukaddam K, Pignolo RJ, et al. Prevalence and risk factors for kidney stones in fibrodysplasia ossificans progressiva. Bone. (2018) 109:120–3. doi: 10.1016/j.bone.2017.12.010
PubMed Abstruse | CrossRef Full Text | Google Scholar
fifty. Sim FJ, Zhao C, Penderis J, Franklin RJ. The age-related decrease in CNS remyelination efficiency is attributable to an impairment of both oligodendrocyte progenitor recruitment and differentiation. J Neurosci. (2002) 22:2451–nine. doi: x.1523/JNEUROSCI.22-07-02451.2002
PubMed Abstract | CrossRef Full Text | Google Scholar
51. Meyers EA, Gobeske KT, Bond AM, Jarrett JC, Peng CY, Kessler JA. Increased bone morphogenetic protein signaling contributes to age-related declines in neurogenesis and noesis. Neurobiol Aging. (2016) 38:164–75. doi: 10.1016/j.neurobiolaging.2015.10.035
PubMed Abstract | CrossRef Full Text | Google Scholar
52. Brady JJ, Li Chiliad, Suthram S, Jiang H, Wong WH, Blau HM. Early role for IL-6 signalling during generation of induced pluripotent stem cells revealed by heterokaryon RNA-Seq. Nat Prison cell Biol. (2013) 15:1244–52. doi: 10.1038/ncb2835
PubMed Abstruse | CrossRef Full Text | Google Scholar
53. Mosteiro L, Pantoja C, de Martino A, Serrano Thousand. Senescence promotes in vivo reprogramming through p16(INK)(4a) and IL-half dozen. Crumbling Cell. (2018) 17:e12711. doi: 10.1111/acel.12711
CrossRef Full Text | Google Scholar
55. Wang H, Lindborg C, Lounev V, Kim JH, McCarrick-Walmsley R, Xu M, et al. Cellular hypoxia promotes heterotopic ossification past amplifying BMP signaling. J Os Miner Res. (2016) 31:1652–65. doi: 10.1002/jbmr.2848
PubMed Abstract | CrossRef Full Text | Google Scholar
56. Freund A, Orjalo AV, Desprez PY, Campisi J. Inflammatory networks during cellular senescence: causes and consequences. Trends Mol Med. (2010) xvi:238–46. doi: x.1016/j.molmed.2010.03.003
PubMed Abstract | CrossRef Full Text | Google Scholar
58. Hayashi Y, Hsiao EC, Sami South, Lancero M, Schlieve CR, Nguyen T, et al. BMP-SMAD-ID promotes reprogramming to pluripotency by inhibiting p16/INK4A-dependent senescence. Proc Natl Acad Sci USA. (2016) 113:13057–62. doi: 10.1073/pnas.1603668113
PubMed Abstract | CrossRef Total Text | Google Scholar
lx. Zhu Y, Tchkonia T, Pirtskhalava T, Gower Ac, Ding H, Giorgadze N, et al. The Achilles' heel of senescent cells: from transcriptome to senolytic drugs. Aging Cell. (2015) xiv:644–58. doi: 10.1111/acel.12344
PubMed Abstract | CrossRef Full Text | Google Scholar
61. Xu Thou, Tchkonia T, Ding H, Ogrodnik K, Lubbers ER, Pirtskhalava T, et al. JAK inhibition alleviates the cellular senescence-associated secretory phenotype and frailty in one-time age. Proc Natl Acad Sci Us. (2015) 112:E6301–10. doi: 10.1073/pnas.1515386112
PubMed Abstruse | CrossRef Full Text | Google Scholar
62. Hino Yard, Horigome K, Nishio M, Komura Southward, Nagata South, Zhao C, et al. Activin-A enhances mTOR signaling to promote aberrant chondrogenesis in fibrodysplasia ossificans progressiva. J Clin Invest. (2017) 127:3339–52. doi: 10.1172/JCI93521
PubMed Abstract | CrossRef Full Text | Google Scholar
schermerhornwilbeend.blogspot.com
Source: https://www.frontiersin.org/articles/10.3389/fendo.2019.00908/full
0 Response to "Bone Book Review Bone Book Review Robert Lubbers"
Postar um comentário